Welcome to HortHerb Publisher
 Harnessing Genetic Populations in Plant Breedi...
Harnessing Genetic Populations in Plant Breedi...- The Fruit Revolution in the Genomic Era: Unvei...
- The Role of Genome-Wide Association Studies (G...
- ECONOMICS OF PRODUCTION AND MARKETING FOR FREN...
- Optimizing Chickpea ( Cicer arietinum L.) Yie...
- Economic Analysis of Apple Production and Mark...
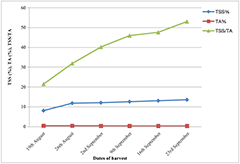
- Effect of Different Postharvest Treatments on ...
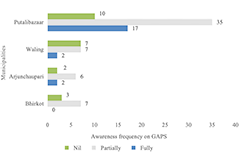
- Productivity Determinants and Production Const...
- A research team from the Chinese Academy of Science...(14 Mar.2024)
- A research team from the Institute of Botany of the ...(14 Mar.2024)
- A research team led by Professor Li Chenlong from S...(01 Mar.2024)
- A research team led by Dr. Zhang Yangyong from the I...(01 Mar.2024)